Uppruni lífsins
—

Uppruni lífsins á jörðinni hefur lengi verið mönnum ráðgáta og hafa ýmsar tilgátur og/eða kenningar komið fram, sem má skipta í 4 meginflokka:
1. Sköpunartilgátur trúarbragðanna.
2. Sjálfskviknunartilgátur um að lífið kvikni af sjálfu sér við réttar aðstæður. Þessar tilgátur liðu að mestu undir lok á 19. öld.
3. Tilgátur um tilflutning lífs til jarðar utan úr geimnum.
4. Efnaþróunarkenningar um að lífið hafi orðið til við efnaþróun.
Snemma á 20. öld fóru menn að velta fyrir sér efnaþróunarmöguleikanum í fullri alvöru. Rússinn Oparin skrifaði bók um uppruna lífsins á fyrri hluta aldarinnar þar sem hann reifaði málið í hnotskurn og ræddi m.a. hugmyndir manna á þeim tíma um efnasamsetningu lofthjúps jarðar og frumhafanna áður en lífið varð til. Árið 1951 gerði Stanley Miller fræga tilraun þar sem hann blandaði saman þeim efnasamböndum, sem menn töldu þá að hefðu verið til staðar á frumjörðinni, hitaði blönduna upp og leiddi í gegnum hana rafneista til að líkja eftir aðstæðum, sem líklegastar þóttu að hefðu verið á jörðinni áður en lífið varð til. Síðan leiddi hann gufuna í gegnum eimsvala, safnaði vökvanum saman og efnagreindi hann. Og viti menn! Í þessum vökva fann hann einfaldar sykrur, stuttar fitusýrur og meira að segja amínósýrur og niturbasa, en þessi efni eru hráefni til byggingar stórsameinda frumu, þ.e. fjölsykra, fituefna, próteina og kjarnsýra. Tilraun Millers hefur verið margendurtekin. Með breyttri og bættri tækni og þekkingu hefur mönnum tekist að fá fram stórsameindir úr ofangreindum 4 efnaflokkum og í rauninni vantar ekkert upp á annað en að fá þessar sameindir til að starfa saman, þ.e. mynda utan um sig himnu, taka inn næringu, láta frá sér úrgang og fjölga sér, en þá hefur okkur tekist að búa til líf úr ólífrænum efnum.
Margt bendir til að í árdaga sólkerfisins hafi aðstæður til efnaþróunar, sem gat leitt til lífs, komið upp víðar en á jörðinni. Enn sem komið er er varla hægt að segja til um þetta með vissu en hafi svo verið virðist sem lífsskilyrði hafi horfið annars staðar en á jörðinni. Í ljósi þessa er undarlegt að meðal sumra stjörnufræðinga eigi tilgátur úr flokki 3 hér að ofan vaxandi fylgi að fagna, og horfa þeir helst til Mars í því sambandi. Þessar tilgátur eiga það sammerkt að lífið hafi orðið til annars staðar en á jörðinni, borist hingað og dáið út þar sem það varð til í upphafi. Ganga sumir jafnvel svo langt að segja að finnist menjar um líf á Mars sé það sönnun þess að lífið á jörðinni sé upprunnið á Mars.
Það er sameiginlegt þessum tilgátum að þær svara í raun ekki spurningunni um hvernig lífið varð til. Þær fara eins og köttur í kringum heitan graut og koma sér hjá því að svara meginspurningunni. Finnist menjar um líf á Mars er það miklu fremur vísbending um að skilyrði til efnaþróunar, sem gat leitt til lífs, hafi komið upp annars staðar í sólkerfinu en eingöngu á jörðinni. Í þessu sambandi má benda á að mönnum hefur tekist að finna sólkerfi, þ.e. sólir með reikistjörnum, utan við okkar sólkerfi. Ef kenningar um tilflutning lífs væru réttar væri eins hægt að segja að lífið hafi orðið til í öðru sólkerfi en borist til okkar sólkerfis um óravíddir himingeimsins. Miklu líklegra er að sú staðreynd að til eru fleiri sólkerfi en okkar bendi til að aðstæður til lífefnaþróunar komi upp í fleiri sólkerfum en okkar.
Ein af stærstu spurningunum varðandi tilurð lífsins, þ.e. skipulagðrar frumustarfsemi, er samstarf próteina og kjarnsýra. Strax árið 1900 voru litningarnir uppgötvaðir. Árið 1928 var staðfest að erfðaefnið væri í litningunum (Griffith o.fl.) og 1944 var sýnt fram á (Avery o.fl.) að DNA væri erfðaefnið. Bygging DNA varð ljós árið 1953 (Watson og Crick) og erfðatáknmálið hefur verið þekkt í áratugi. Nú er það svo að DNA geymir upplýsingar um myndun próteina, þ.m.t. ensíma, sem eru sérhæfð prótein, sem stýra og hraða nánast öllum efnahvörfum í lífverum. Enn fremur flytjast þessar upplýsingar milli kynslóða með DNA, sem getur myndað nákvæma eftirmynd af sjálfu sér. Til að DNA geti eftirmyndað sjálft sig og stjórnað ensímmyndun þarf ensím og er þetta stærsta mótsögn sameindalíffræðinnar. Hvort var á undan, eggið eða hænan? Voru það kjarnsýrurnar, sem yfirtóku próteinin og gerðu þau að þrælum sínum eða voru það próteinin sem yfirtóku kjarnsýrurnar og gerðu þær að gagnabanka?
Dr. Guðmundur Eggertsson, fyrrverandi prófessor í erfðafræði við Háskóla Íslands, hefur velt þessu talsvert fyrir sér og skrifað um þetta nokkrar bækur. Við skulum aðeins glugga í niðurstöður hans.
Eins og sagði hér að ofan getur efnaþróun leitt til myndunar hráefna til byggingar stórsameinda í frumum eins og við þekkjum þær á jörðinni. Hins vegar eru einstakir byggingahlutar (amínósýrur í próteinum og niturbasar í kjarnsýrum) gersamlega óvirkir nema sem hluti af viðkomandi stórsameindum. Því er vert að skoða líklegustu efnaþróun stórsameinda miðað við þekkingu okkar á tilurð þeirra í árdaga jarðar.
Til að byrja með er mjög mikilvægt að gera sér grein fyrir að á þeim tíma var ekkert óbundið súrefni í lofthjúpnum. Súrefni hefur þann skrýtna eiginleika að ráðast á alls konar efnatengi og efnahópa í lífrænum efnasamböndum og breyta þeim eða jafnvel brjóta niður. Þessir efnaferlar eru oft kallaðir oxun (súrefni heitir oxygen á mörgum erlendum málum) og geta eingöngu átt sér stað þar sem súrefni er til staðar og á greiða leið að efnatengjunum eða hópunum. Öfugt við oxun eru svokölluð afoxunarefnahvörf og má segja að lífræn efnasambönd endist lengur í afoxandi umhverfi.
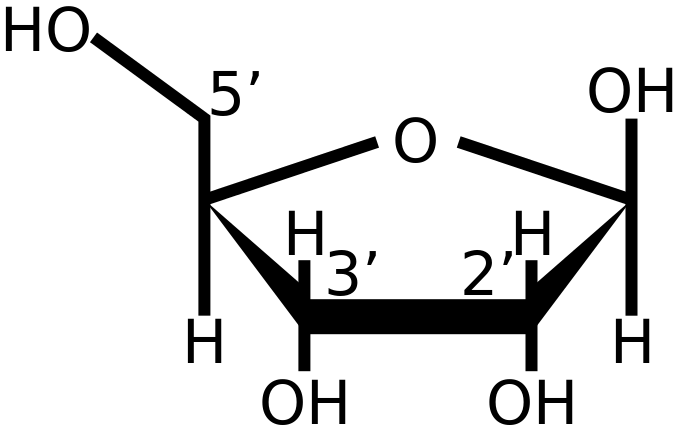
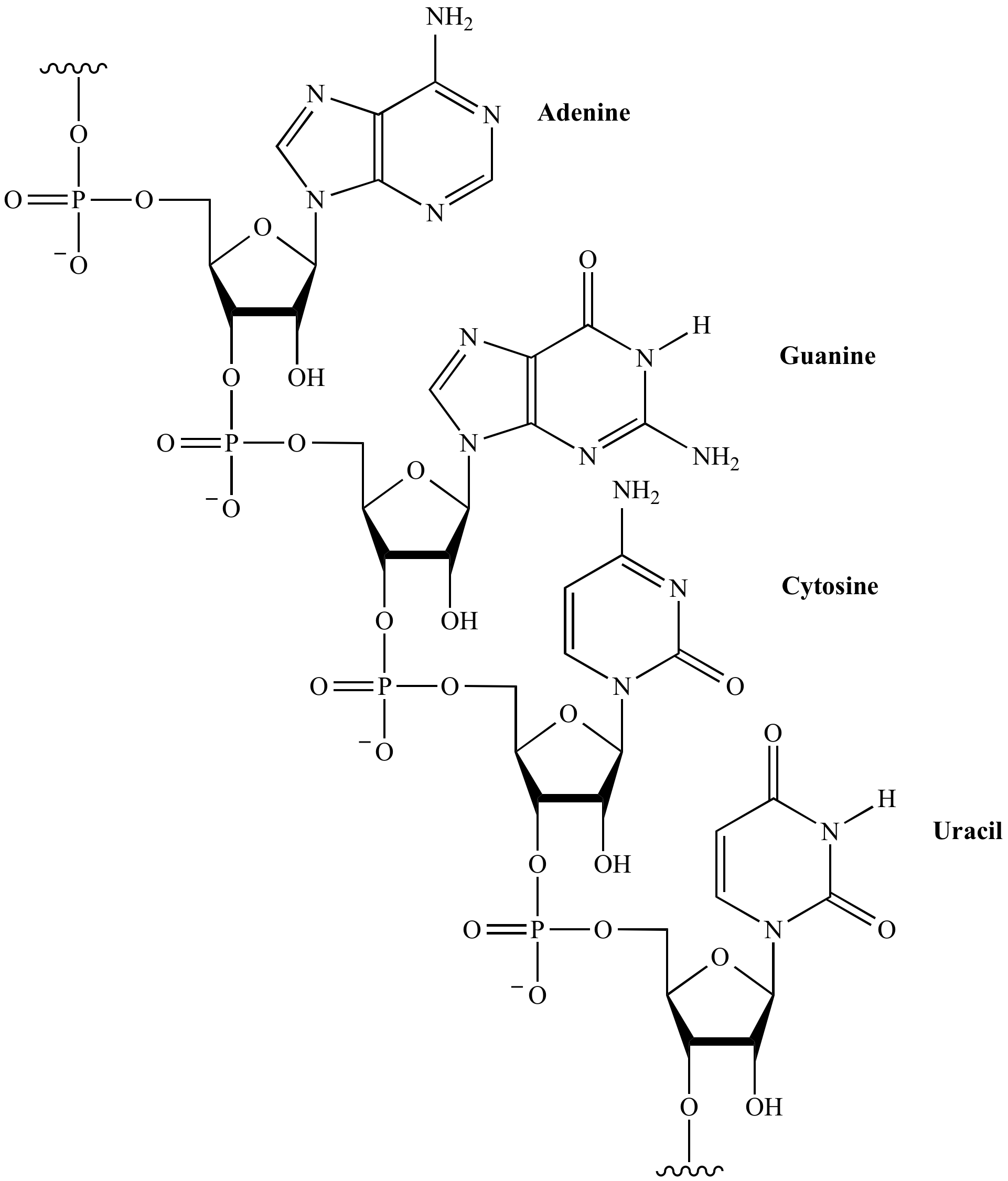
Nú, ef lofthjúpurinn var súrefnislaus í upphafi og þar með afoxandi, er líklegt að kjarnsýran RNA (riboxy nucleic acid eða ríbósakjarnsýra) hafi orðið til við samruna svokallaðra kjarnsýrueininga (nucleotíða) langt á undan DNA (deoxyribose nucleid acid) einmitt vegna þess að RNA er mjög viðkvæm fyrir súrefni en þolir afoxandi umhverfi mjög vel. RNA-einingar (nucleotíð) samanstanda af sykrunni ríbósa, fosfatsameind og einum af fjórum mögulegum niturbösum; adenín, gúanín, cytosín og uracil, þ.e. fosfat-ríbósi-niturbasi. Ríbósinn er sykra úr 5 kolefnisatómum og er með svokallaðan OH hóp eða hydroxyl hóp á kolefni tvö (sjá mynd). Kolefni nr. 1 (hægra megin) heldur á niturbasanum og kolefni 3 heldur á fosfathópnum. Þetta þýðir að OH hópurinn á kolefni 2 er sá eini, sem er viðkvæmur fyrir súrefnisárásum þar sem hinir hóparnir eru uppteknir við annað þar sem kolefni 5 á einu nucleotíði tengist fosfati á kolefni 3 á næsta nucleotíði og þannig myndast RNA keðjan (sjá mynd). Þannig eru allar RNA keðjur meira eða minna opnar og með óvarinn OH hóp á kolefni 2 á hverjum einasta ríbósa í keðjunni.
Nú, samt sem áður er RNA sérlega duglegt að raða utan á sig amínósýrum, byggingarefnum próteina, og raðast þær á niturbasana og þarf þrjá niturbasa í röð til að raða á sig einni amínósýru. Þetta bendir eindregið til þess að RNA hafi verið upprunalegi lykillinn að próteinmyndun á jörðinni og enn fremur verið forveri DNA, sem er nokkuð flóknari sameind.
DNA er skammstöfun fyrir 2-Deoxyribose Nucleic Acid (deoxykjarnsýra). Það þýðir að OH hópurinn á kolefni 2 í sykursameindinni hefur verið afoxaður og því heitir sameindin ekki lengur ríbósi, heldur 2-deoxyríbósi (sjá mynd). Þetta þýðir að það er enginn hópur lengur, sem er viðkvæmur fyrir súrefnisárásum, sem er að sjálfsögðu mjög hagstætt í súrefnisríku umhverfi. Auk þess er DNA tvöfaldur þráður en ekki einfaldur (sjá mynd) sem aftur veitir enn meiri vörn gegn efnaárásum.
Þannig er nú talið fullvíst að þróunarröð stórsameindanna hafi verið RNA – prótein – DNA. Allar lífverur á jörðinni nota DNA sem erfðaefni, þ.e. sem geymslu erfðaupplýsinga um próteinframleiðslu, sem flyst á milli kynslóða. Til eru svokallaðar RNA-veirur, sem geyma erfðaupplýsingar á RNA formi en veirur eru strangt tekið ekki lífverur. DNA er sem sagt efnafræðilega mun heppilegra erfðaefni en RNA og kannski má segja að það hafi yfirtekið geymslu- og miðlunarhlutverkið en RNA er samt sem áður boðberinn milli niturbasanna og amínósýranna í próteinunum. M.ö.o. miðlar RNA erfðaupplýsingunum frá DNA til próteinanna. Leið erfðaboðanna í öllum frumum með erfðaefni er því DNA – RNA – prótein og það er engin leið til baka.














{kind=link}
{kind=link}
{kind=link}